第五届中国出版政府奖音像电子网络出版物奖提名奖
中国科技核心期刊
中国科学引文数据库(CSCD)来源期刊

第五届中国出版政府奖音像电子网络出版物奖提名奖
中国科技核心期刊
中国科学引文数据库(CSCD)来源期刊
2024 , Vol. 10 >Issue 03: 279 - 286
DOI: https://doi.org/10.3877/cma.j.issn.2096-1537.2024.03.012
单细胞测序技术在脓毒症免疫研究中的应用进展
Copy editor: 卫轲
收稿日期: 2024-04-30
网络出版日期: 2024-09-06
基金资助
宁夏回族自治区重点研发计划项目(2023BEG02024)
版权
Application of single cell sequencing technology in the study of sepsis immunity
Received date: 2024-04-30
Online published: 2024-09-06
Copyright
苏生林 , 马金兰 , 于弘明 , 杨晓军 . 单细胞测序技术在脓毒症免疫研究中的应用进展[J]. 中华重症医学电子杂志, 2024 , 10(03) : 279 -286 . DOI: 10.3877/cma.j.issn.2096-1537.2024.03.012
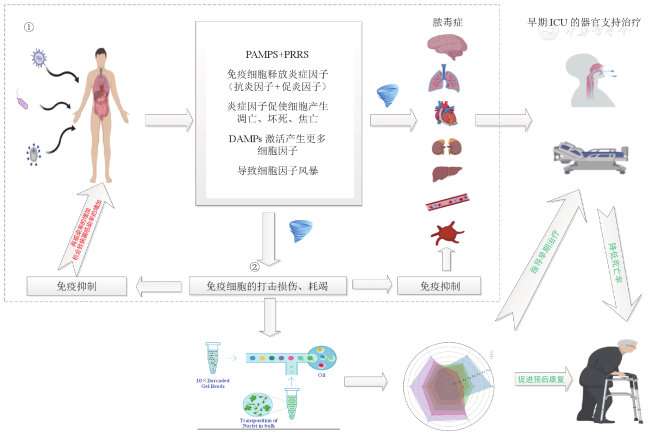
Sepsis is a clinical syndrome with a high mortality rate, and despite the development and advancement of multidisciplinary approaches, sepsis can progress to fatal organ failure. In recent years, the rapid development of single-cell sequencing technology has provided a new technical support for the exploration of the pathophysiological mechanism of sepsis, especially the application in immunology has made the precise prevention and treatment of sepsis possible. This review reviewed relevant studies in recent years, described the occurrence and development of sepsis and sepsis-induced immunosuppression, introduced the application progress of single cell sequencing technology in sepsis and sepsis-induced immunosuppression research, and aimed to explore the possibility of future application of single cell sequencing technology in the precision treatment of sepsis.
Key words: Sepsis; Single-cell RNA sequencing; Immunosuppression; Precision medicine
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}