第五届中国出版政府奖音像电子网络出版物奖提名奖
中国科技核心期刊
中国科学引文数据库(CSCD)来源期刊

第五届中国出版政府奖音像电子网络出版物奖提名奖
中国科技核心期刊
中国科学引文数据库(CSCD)来源期刊
2026 , Vol. 12 >Issue 01: 93 - 100
DOI: https://doi.org/10.3877/cma.j.issn.2096-1537.2026.01.014
蛋白质-合成代谢抵抗导致老年脓毒症患者骨骼肌质量下降机制的研究进展
通信作者:
丁欢,Email:iresrainbow@sina.comCopy editor: 卫轲
收稿日期: 2024-09-21
网络出版日期: 2026-04-29
基金资助
宁夏回族自治区重点研发计划项目(2022BEG03125)
宁夏回族自治区科学技术厅自然科学基金项目(2024AAC03544)
版权
Research progress on the mechanism of protein-anabolic resistance leading to decreased skeletal muscle mass in elderly sepsis
Corresponding author:
Ding Huan, Email: iresrainbow@sina.comReceived date: 2024-09-21
Online published: 2026-04-29
Copyright
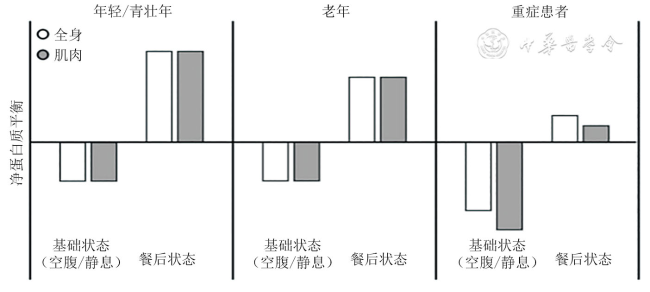
骨骼肌质量下降是脓毒症严重并发症之一,其特征是在组织学水平上肌力和纤维横截面积减弱、缩小和减少。表现为四肢松弛无力,尤其是近端肢体肌肉,同时会影响患者特别是老年患者的预后,降低生活质量,危及生命安全。脓毒症患者合成代谢抵抗导致蛋白质合成和分解之间的平衡被破坏,表现为蛋白质分解代谢增加与合成代谢减少,是造成骨骼肌质量下降的主要原因之一。目前,脓毒症骨骼肌分解代谢相关研究比较明确,蛋白质-合成代谢抵抗成为日趋关注的问题。因此,本文就蛋白质-合成代谢抵抗与脓毒症骨骼肌质量下降的相关性机制进行综述,为脓毒症骨骼肌质量下降的临床防治提供理论依据和方案。
关键词: 老年脓毒症; 骨骼肌质量; 蛋白质-合成代谢抵抗
李锦源 , 麦涵钰 , 陈凯 , 周文艳 , 白吉佳 , 曹相原 , 丁欢 . 蛋白质-合成代谢抵抗导致老年脓毒症患者骨骼肌质量下降机制的研究进展[J]. 中华重症医学电子杂志, 2026 , 12(01) : 93 -100 . DOI: 10.3877/cma.j.issn.2096-1537.2026.01.014
The decline of skeletal muscle mass is one of the serious complications of sepsis. It is characterized by the weakend muscle strength, reduced fiber cross-sectional area and dismished muscle quantity. It manifests as flaccid weakness in the limbs, particularly in the proximal limb muscles. Meanwhile, it will affect the prognosis of patients, especially elderly patients, leading to their reduced quality of life and life safety risk. The anabolic resistance in sepsis patients leads to the disruption of the balance between protein synthesis and breakdown, manifested as increased protein catabolism and decreased anabolism, which is one of the main reasons for the decline of skeletal muscle mass. At present, the relevant research on skeletal muscle catabolism in sepsis is relatively well estabished, and protein-anabolic resistance has become an issue of increasing concern. Therefore, we review the correlative mechanisms between protein-anabolic resistance and the decline of skeletal muscle mass in sepsis, aiming to provide theoretical basis and solutions for the clinical prevention and treatment of the skeletal muscle loss in sepsis patients.
李锦源, 麦涵钰, 陈凯, 等. 蛋白质-合成代谢抵抗导致老年脓毒症患者骨骼肌质量下降机制的研究进展[J/OL]. 中华重症医学电子杂志, 2026, 12(1): 93-100.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}